Núcleo, retículo endoplasmático, aparato de Golgi y ribosomas

Núcleo. Excepto los tubos cribosos
(liberianos) en estado adulto, todas las células vegetales presentan núcleo. En
las meristemáticas, el núcleo es de gran tamaño; en las células del embrión
pueden ocupar más del diámetro de la cavidad celular; en tanto que en las más
diferenciadas el núcleo es menor.
En estado
interfásico (de reposo), el núcleo presenta una estructura definida que se
pierde al iniciarse la mitosis (forma de reproducción celular). En la interfase
el núcleo tiene forma esférica aunque puede variar a elipsoidal, discoidal o
alargada. Está limitado por una envoltura doble constituida por lípidos y
proteínas insolubles; y por estudios hechos con el microscopio electrónico, la
membrana externa presenta una serie de comunicaciones con el retículo endoplasmático.
Por dentro de la envoltura nuclear existe una matriz llamada jugo nuclear o
cariolinfa, en la que se encuentran incluidos la cromatina y uno o más
nucléolos. La cromatina es un compuesto reticular constituido por proteínas y
ADN; durante la división celular, la cromatina se organiza para formar los
cromosomas que transfieren la información genética; el número y la
configuración de estos es constante para cada especie. Los cromosomas son
visibles sólo cuando el núcleo se halla en estado de división. Los
nucléolos son cuerpos densos y más o menos homogéneos. El número de nucléolos
por núcleo es generalmente de uno, pero puede haber dos o más, rara vez están
ausentes. Durante la división celular, el núcleo y los nucléolos desaparecen
por completo y vuelven a aparecer cuando concluye el proceso de división
celular.
El núcleo
efectúa funciones vitales para la célula tales como: la síntesis proteica,
control de las funciones celulares y dependiendo del código genético y durante
la división celular, la transmisión de los caracteres hereditarios. La remoción
del núcleo va seguida de la muerte de la célula.

Retículo
endoplasmático.
Aunque en un principio se consideró al protoplasma como un coloide homogéneo.
Después se descubrió que estaba atravesado por una serie de membranas dobles
(pares de membranas) a manera de túbulos simples o ramificados que pueden estar
comunicados con la membrana plasmática y la nuclear; a esta estructura se le
denominó retículo endoplasmático. Las membranas pueden plegarse y constituir
cisternas o vesículas y en ocasiones pueden continuarse entre una célula y otra
a través de los plasmodesmos. La naturaleza química de estas membranas es de
lípidos y proteínas. El retículo puede o no estar asociado con los ribosomas,
en el primer caso se le llama retículo rugoso y en el segundo, retículo liso.
Al retículo se le atribuyen las siguientes funciones: transporte, y
almacenamiento de proteínas, interviene en la formación de la pared celular y
junto con el citoplasma interviene en la formación de ácidos grasos.

Aparato de
Golgi (dictiosoma). En las
células animales, los dictiosomas forman el aparato de Golgi. En 1956 fueron
reconocidas estas estructuras en las células vegetales, ya que en éstas, no se
encuentra el cuerpo de Golgi como en las células animales, organizado como un
complejo cerca del núcleo, sino que están libres en el citoplasma. Los
dictiosomas están constituidos por cisternas o discos, formados por membranas
dobles y cercanas entre sí. En los vegetales, los dictiosomas sintetizan
material que se utiliza en la pared celular, para ello, se desprenden de las
cisternas, vesículas conteniendo el material sintetizado, las cuales emigran
hacia la pared donde descargan su contenido, y se fusionan con ésta, y de la
misma forma actúan en la telofase para formación de la placa celular.
Ribosomas
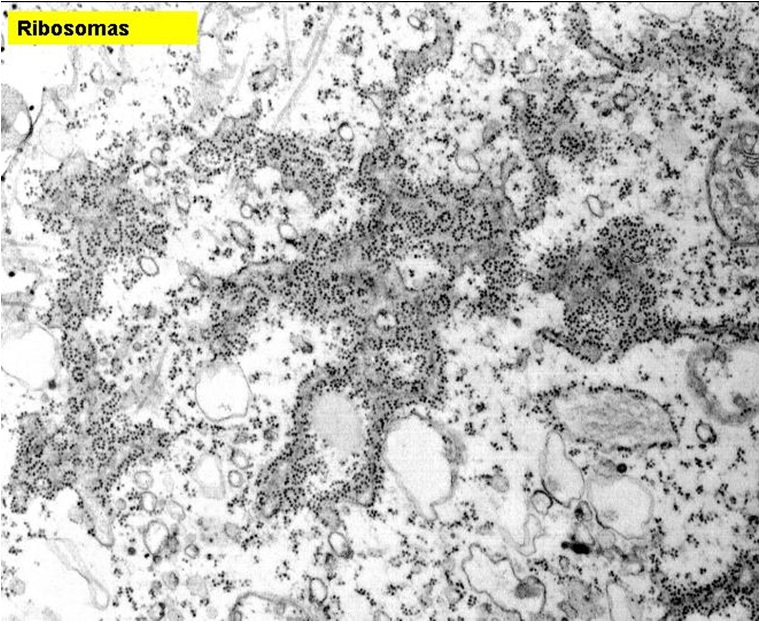 |
| Gránulos compuestos de ARN y proteínas; algunos unidos al retículo endoplasmático rugoso, otros libres en el citoplasma. |
Los
ribosomas son organelos que se encuentran en todas las células, tanto
procariotas como eucariotas. Se encargan de la síntesis proteínica. Estos
organelos se pueden apeciar como pequeños gránulos opacos al paso de electrones
(en microscopía electrónica), que se asocian a la matriz externa del retículo
endoplásmico (rugoso). A estos gránulos se le ha llamado de Palade porque fue
este investigador quien los descubrió en 1955; ahora el nombre más aceptado es
el de ribosomas. Miden
aproximadamente 10 nm y están constituidos por ribonucleoproteína.

Brachet y
Casperon fueron los primeros (1941) en sugerir que las células ricas en ARN
están relacionadas con la síntesis de proteínas. Por ejemplo, en los
reticulocitos en estado de diferenciación (es decir, eritrocitos de mamífero,
pero en estado inmaduro) se presenta una disminución tanto en el contenido de
ácido ribonucleico (ARN), como en la velocidad de incorporación de aminoácidos
a la molécula proteínica; en cambio, en culivos de bacterias en crecimiento
sucede todo lo contrario. Es así que en Escherichia coli se ha podido calcular
que existen alrededor de 10000 ribosomas por célula. Los ribosomas en este caso
se hallan en forma libre, como sucede en toda célula procariótica que no posee
retículo endoplásmico.
Los
ribosomas tienen una estructura sumamente compleja, por ejemplo, los ribosomas
de E. coli denominados 70S están constituidos por dos subunidades que
pueden ser separadas; una es mayor, de 50S y otras más pequeña, de 30S (el
término S se refiere al tamaño que se determina por la velocidad de
sedimentación medida en unidades Svedberg, o sea, la velocidad a la cual una
partícula sedimenta cuando es centrifugada). El ribosoma completo (70S) tiene
un peso de alrededor de 2.7 millones de Daltones y contiene 65% de ARN y 35% de
proteínas.
Las
subunidades pequeñas (30S) contienen a su vez una molécula de ARN de 16S y 21
diferentes proteínas ribosomales. La subunidad de 50S contiene por su parte un
ARN de 5S y otro ARN con un valor de 23S, además de 34 diferentes moléculas de
proteínas ribosomales.
Los
ribosomas de organismos eucariotes son esencialmente idénticos a los de E.
coli, aunque difieren en el tamaño de la subunidades y en el número de
proteínas ribosomales.
 |
| Polisomas o polirribosomas vistos con el microscopio electrónico. Los de (a) provienen de reticulocitos de conejo y están ocupados en la traducción de ARNm de la hemoglobina y los de (b) proviene de células de glándulas salivares de la mosca de agua Chironomus thummi. En (b) se puede ver como emerge la cadena polipeptídica naciente de cada ribosoma. Su longitud incrementa al avanzar la traducción, desde la izquierda (5’) a la derecha (3’) del ARNm. |
Cabe
mencionar, también, que la función biológica de los ribosomas es altamente
compleja, ya que involucra el reconocimiento y la capacidad de unirse al ARN
mensajero formando los polisomas, y a los ARN de transferencia específicos para
aminoácidos; intervienen en la formación de los enlaces peptídicos y en su
traslocación física a lo largo del ARN mensajero. En este sentido se ha
logrado, gracias a las investigaciones en especial de M. Nomura y sus
colaboradores, el poder delinear varias de las principales características del
autoensamble de los ribosomas y la secuencia necesaria en los procesos de
ensamblado. Por medio de estas investigaciones se ha llegado a saber que cuando
se exponen las subunidades 30S de los ribosomas de E. coli a altas concentraciones de sal, se origina la pérdida de
siete distintas proteínas ribosomales, dejando un "núcleo" con un
valor de 23S que consiste de ARN y proteína. Puede observarse, entonces, que
esos "núcleos" de ribosoma ya no tienen actividad en la síntesis de
proteínas.
De las 7 proteínas solubilizadas se han podido purificar cinco y las otras dos se han obtenido como mezcla. Se sabe que las cinco proteínas puras tienen diferente composición en aminoácidos. Con los resultados anteriores ahora es posible reconstituir ribosomas activos en la síntesis de proteínas al mezclar el "núcleo" de 23S con las proteínas previamente solubilizadas, ya que al añadir a esta mezcla las subunidades ribosomales de 50S se obtiene un material capaz de sintetizar una cadena polipeptídica, pero, por supuesto, se deben añadir los componentes esenciales para la síntesis de proteínas. Sin embargo, cuando se elimina cualquiera de las cinco proteínas purificadas, la síntesis tiende a disminuir en forma drástica.
De esta forma, y con otros experimentos de tipo genético tendientes a conocer las proteínas que constituyen los ribosomas se ha podido llegar a entender cuál es la función que desempeñan dichas proteínas como distintos componentes ribosomales en la síntesis de proteínas.
Recientemente, se ha logrado avanzar también en lo que se refiere a la reconstitución de los ribosomas con las subunidades de 50S.
El significado de los resultados de ensamble obtenidos a partir de las subunidades de 30S puede resumirse en lo siguiente:
1. Toda información necesaria para el ensamble correcto está en la estructura de los compuestos de este organelo y no se requieren otros factores. O sea que el proceso tanto in vitro como posiblemente in vivo es de autoensamble.
2. Se puede asegurar por medio de estos experimentos de reconstitución cuándo un compuesto es esencial para el ensamble del ribosoma, o para una función determinada. En este sentido se ha podido identificar el componente responsable para la sensibilidad al antibiótico estreptomicina que inhibe la iniciación en la síntesis de proteína.
Fuente: González Embarcadero, Antonia; Cedillo Portugal, Ernestina; Díaz Garduño, Lucina. 2011. Morfologiía y anatomía de las plantas con flores. Universidad Autónoma Chapingo. México.



No hay comentarios.:
Publicar un comentario